Article Text

Abstract
Background The well-known ‘pyrotherapy’ of Julius Wagner-Jauregg might be the beginning of the study on the immunological concepts of schizophrenia. As the primary immune effector cells in the brain, microglia play a pivotal role in neuroinflammatory processes. Maternal viral infection during pregnancy is associated with an increased risk for psychiatric disorders with presumed neurodevelopmental origin, including autism spectrum disorders and schizophrenia. The present study was to quantify microglia activation in vivo in the mature offspring of rats exposed to polyriboinosinic–polyribocytidilicacid (Poly I:C) during pregnancy using 11C-PK11195 positron emission tomography (PET) and immunohistochemistry.
Objective The study aimed to quantify microglia activation in vivo in the prefrontal cortex and hippocampus in mature offspring of prenatal Poly I:C exposed rats.
Methods Offspring of Poly I:C-treated dams were the model group, offspring of saline-treated dams were the control group. Behavioural test for two groups was taken by spontaneous activity, prepulse inhibition (PPI) and latent inhibition (LI) test (including active avoidance conditioning task and passive avoidance conditioning task). Randomly selected successful model rats were assessed by behavioural test in the model group and control group rats. 11C-PK11195 micro-PET/CT and immunohistochemistry were performed on the selected rats to measure microglia activation.
Results The treatment group showed hyperlocomotion and deficits in PPI and LI compared with the control group. The treatment group also showed an increased 11C-PK11195 uptake ratio in the prefrontal cortex (t=−3.990, p=0.003) and hippocampus (t=−4.462, p=0.001). The number of activated microglia cells was significantly higher in the treatment group than in the control group (hippocampus: t=8.204, p<0.001; prefrontal: t=6.995, p<0.001). Within the treatment group, there were significant correlations between the behavioural parameters and the activation of microglia as measured by PET and immunohistochemistry.
Conclusions The present study demonstrated microglia activation in vivo in the prefrontal cortex and hippocampus in mature offspring of prenatal Poly I:C exposed rats. This study suggests that microglia activation may play a possible or potential role in the pathogenesis of schizophrenia.
- schizophrenia
- PET
- neuroinflammation
- microglia
- hippocampus
This is an Open Access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0
Statistics from Altmetric.com
Introduction
The well-known ‘pyrotherapy’ of Julius Wagner-Jauregg might be the beginning of the study on the immunological concepts of schizophrenia (SCZ). Our previous studies reported elevated serum levels of interleukin-1β (IL-1β) and tumour necrosis factor (TNF)-α (TNF-α), increased nuclear factor-кB (NF-κB) activation and its mRNA expression, and activated Th17 cells in peripheral blood mononuclear cells in patients with first-episode SCZ.1 A human postmortem study has shown increased cytokine levels in the brains of patients with SCZ.2 As the primary immune effector cells in the brain, microglia play a pivotal role in neuroinflammatory processes.3
Maternal viral infection during pregnancy is associated with an increased risk of psychiatric disorders with presumed neurodevelopmental origin, including autism spectrum disorders and SCZ.4 Bilbo and colleauges have hypothesised that subsets of microglia are permanently maintained in an activated or primed state into adulthood as a consequence of prenatal infection, and that a subsequent immune challenge in adulthood can cause exaggerated levels of cytokines from these already primed microglia.5 6
Recently, a great deal of interest has been centred on the establishment of neurodevelopmental rodent models in which the basic experimental manipulation takes the form of prenatal exposure to infection and/or immune activation.7 In addition to the influenza infection animal model,8 an animal model has been increasingly used in SCZ research: offspring of female mice that were administered viral mimetic polyriboinosinic–polyribocytidilic acid (Poly I:C) during pregnancy. Those offspring reportedly show changes in brain morphology, physiology, chemistry as well as behaviours that are similar to changes observed in patients with SCZ.9 10 Specifically, prenatal Poly I:C treatment on the gestation date (GD) 9 leads to a pathological profile in the offspring characterised by suppression in exploratory behaviour, abnormalities in selective associative learning in the form of latent inhibition (LI) disruption and abolition of the US pre-exposure effect, impairments in sensor motor gating in the form of reduced prepulse inhibition (PPI), enhanced sensitivity to the indirect dopamine receptor agonist amphetamine and deficiency in spatial working memory when the demand on temporal retention is high.11 12 The findings from our group showed that the serum levels of cytokines (TNF-α and IL-10) in the offspring increased after an injection of Poly I:C in rats during early pregnancy, and the NF-кB inhibitor pyrrolidine dithiocarbamate suppressed the maternal immune response induced by Poly I:C and partially prevented the behavioural changes in the offspring that mimicked neurodevelopmental disorders.
It is well known that microglia activation can be visualised in vivo using 11C-PK11195 (PK11195) positron emission tomography (PET).13 14 The present study was to quantify microglia activation in vivo in the mature offspring of rats exposed to Poly I:C during pregnancy using 11C-PK11195 PET and immunohistochemistry. The brain regions of interest (ROI) included the prefrontal cortex and hippocampus, which have been frequently reported to be involved in the pathogenesis of SCZ.
Methods and materials
Animals
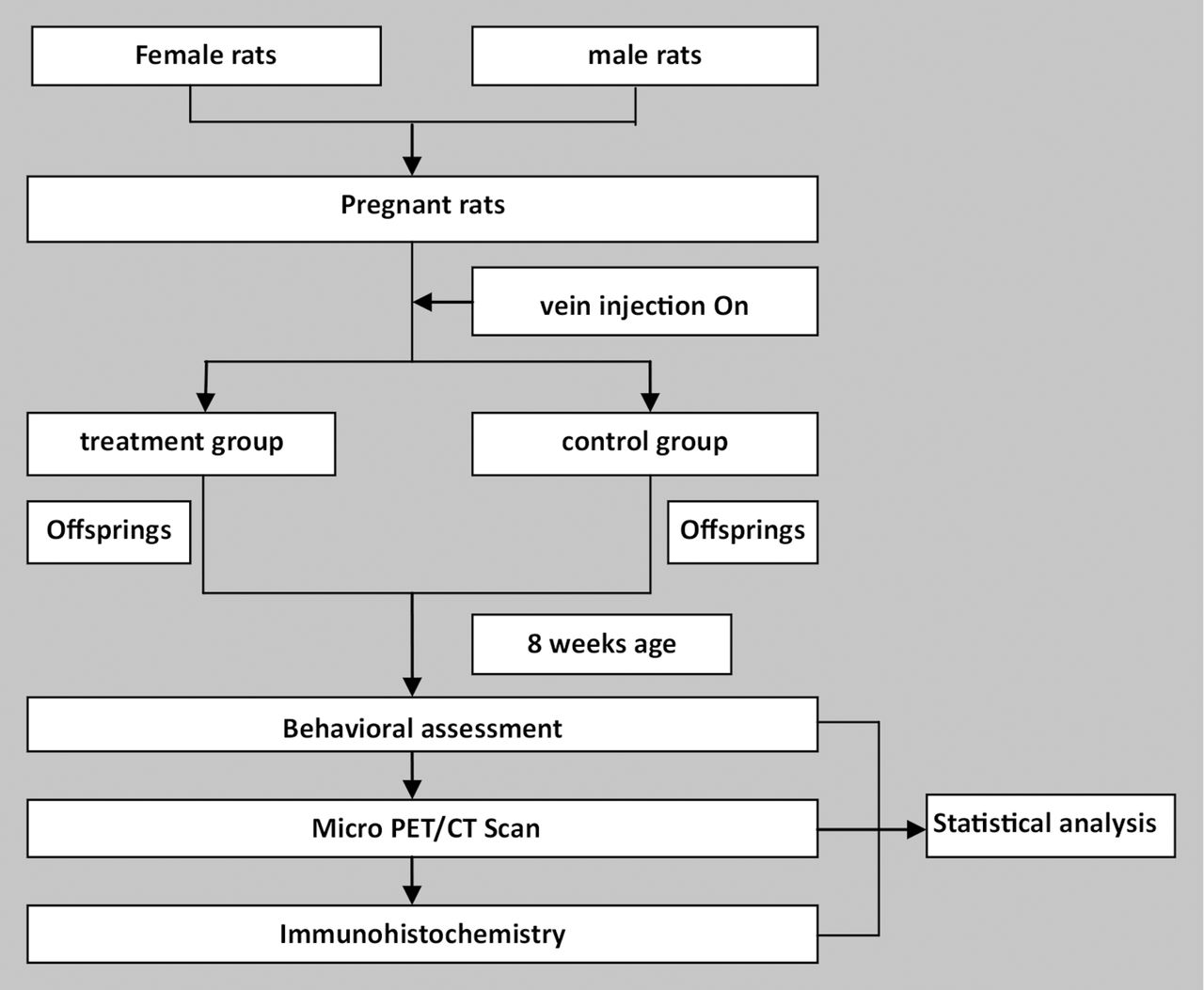
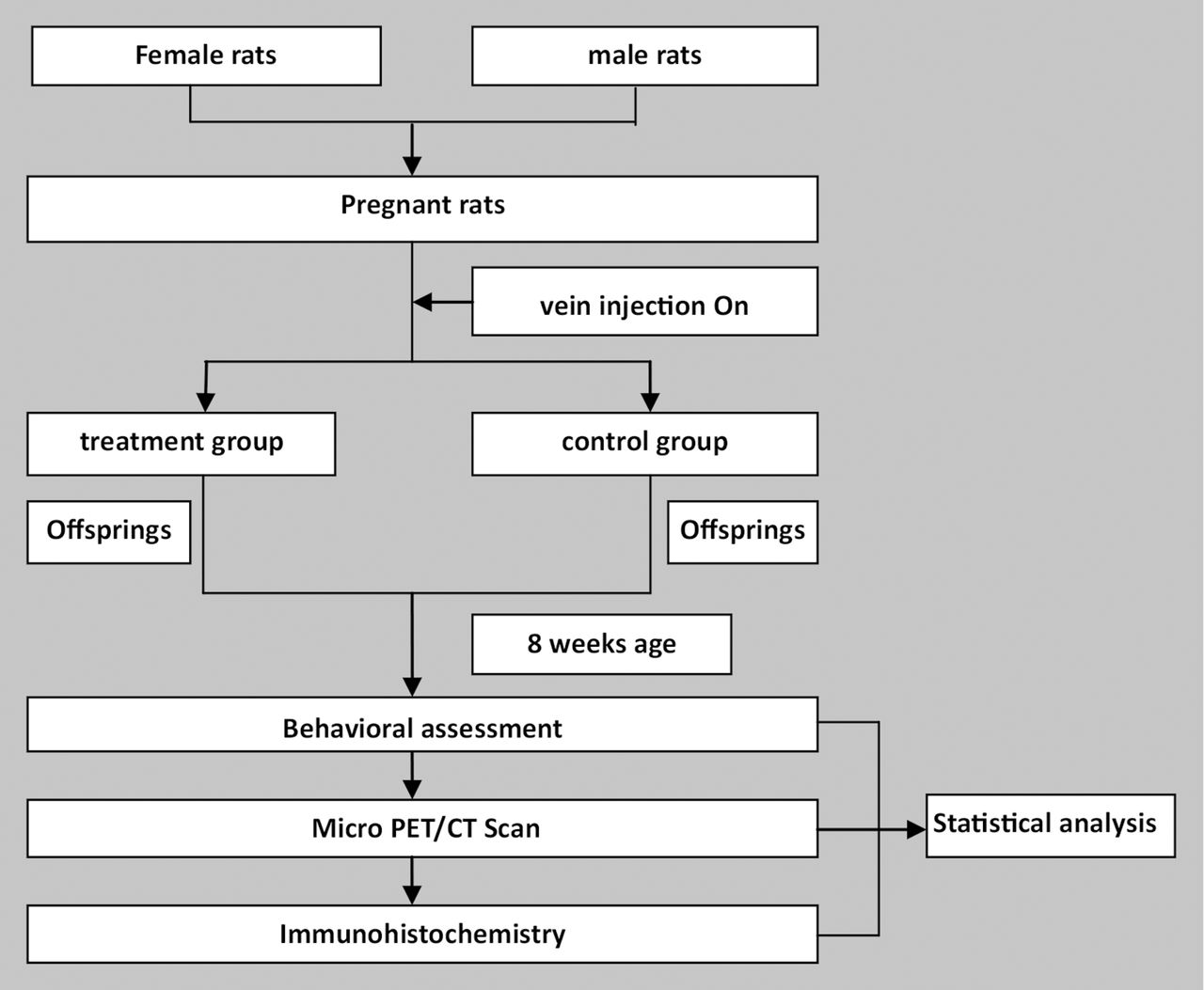
Female and male-specific pathogen-free colony Sprague-Dawley (SD) rats (10–12 weeks old) were obtained from the Laboratory Animal Center of Zhengzhou University (Zhengzhou, Henan, China) and individually housed in a temperature and humidity-controlled environment with a 12-hour light/12-hour dark cycle (lights on at 07:00). Food and tap water were available ad libitum. All animal experiments in the present study were approved by the Animal Care and Use Committee at Xinxiang Medical University, Xinxiang, China, where the experiment was carried out. After 2 weeks of acclimatisation, male and female rats were colocated overnight. Male rats were removed in the morning of the next day. The breeding was stopped until 18 pregnant rats were accomplished. The successful copulation verified by the presence of a vaginal plug was considered GD 1.
Modelling method
Eighteen pregnant rats were randomly divided into two groups. On GD 9, all pregnant dams were weighed, and received a single caudal vein injection of Poly I:C or saline. The treatment group was defined as the offspring of pregnant dams (n=9) that received a 10 mg/kg Poly I:C injection15 16; the control group included the offspring of pregnant dams (n=9) that received a 10 mg/kg saline injection. The injections were performed between 09:00 and 10:00 to minimise the variability associated with diurnal rhythms.
Behavioural assessment
The offspring were weaned at postnatal day 21. Four offspring were housed to a cage by sex and litter, and maintained undisturbed until 8 weeks of age. The gender ratio of the treatment group and control group was approximately 1:1. One of each littermate was randomly selected for behavioural testing. The number of subjects employed in the behavioural testing is summarised in table 1.
Sample size of each group and the sequence of behavioural testing
Locomotor activity
An open-field test was used to assess locomotor activity in a 100 cm× 100 cm× 30 cm box. Data collection was performed by the SMART video tracking program (Panlab, Spain). To begin testing, the animals were gently placed in the centre of the area, and allowed to explore for 30 min. Rats were tested for 60 min and data were captured.
Latent inhibition
LI included both passive and active avoidance tasks.17 The passive avoidance conditioning task was carried out over 2 days. On the training day, each rat was placed in the lighted compartment, facing away from the dark compartment. After 5 s, the door was lifted. As soon as the rat entered the dark compartment with all four paws, the guillotine door was closed. The latency from the time the door was lifted to the time the rat entered the compartment was recorded as T1. A foot shock (0.5 mA, 2 s duration) was delivered 3 s after the door was closed. On the test day (24 hours after training), the same experiment procedure was followed as on the training day. The latency from the time the door was lifted to the time the rat entered the compartment was recorded as T2. The rat then returned to the home cage. T was calculated as the difference between T1 and T2.
The active avoidance task is a fear-motivated associative avoidance test using an electric current as a source of punishment. In our study, the active avoidance conditioning was carried out in a two-way shuttle box apparatus (Panlab). The subject acclimated to the training box for 10 min before the electric bulb (10 W) (unconditioned stimulus) was turned on in the compartment where the rat was located. Thirty seconds later, an electric current (0.3 mA, 5 s) (conditioned stimulus) was delivered to the grid floor of the compartment. The electric current and the light were turned off simultaneously after the rat was transferred into a different compartment. Each rat received 100 trials per day for five consecutive days.18 The rate of active avoidance conditioned response was calculated as the total number of conditioned responses/500. A higher value indicated better learning and memory.
Prepulse inhibition
PPI is the normal suppression of a startle response when a low-intensity stimulus, which elicits little or no behavioural response, immediately precedes an unexpected stronger startling stimulus. The amount of PPI is expressed as the percentage decrease in the amplitude of the startle response caused by the presentation of the prepulse.
All test sessions were performed in a single chamber startle apparatus (QMC-I, Kunming Institute of Zoology, Chinese Academy of Sciences, China).17 One rat was tested during each experimental session. In the test session, the rats were acclimated in the testing cylinder for 5 min, during which time the rats received background noise of only 70 dB sound pressure level (SPL). The test began with six trials of a pulse-alone startle stimulus, consisting of a 40 ms burst of white noise of 120 dB SPL. The session continued with 20 randomised trials, which included five trials of a pulse-alone stimulus and five trials of each of the three types of prepulse startles. A prepulse startle consisted of a prepulse (white noise at 2, 4 or 8 dB SPL above the 70 dB SPL background), a 100 ms interval and a startle pulse (40 ms, 120 dB SPL white noise). The interval length between the 20 randomised trials varied randomly from 8 to 23 s with an average of 15 s. Startle responses were measured with the program developed by Kunming Institute of Zoology, Chinese Academy of Sciences, China. The amount of PPI was expressed as the percentage decrease in the amplitude of the startle response caused by the presentation of the prepulse. The amplitude of the startle response without a prepulse is p. When a weak stimulus is given prior to the startle reflex stimulus, the amplitude for the startle response is pp. The percentage of PPI for each rat was calculated as (1-pp/p)×100, which is proportional to the inhibitory effect of PPI. Using this description of PPI, a high degree of sensory motor gating is reflected in a high % PPI value, whereas a lower or no gating results in a small or negative % PPI value.
Micro-PET/CT scan
11C-PK11195 was synthesised from the precursors 11C-Triflate-CH3 (ABX Express, Germany) and PK11195, with a radiochemical purity of more than 98%, pharmaceutical specifications: specific activity 62.5 TBq/mol.
Rats were anaesthetised for the duration of the scan session with 5% isoflurane using a system built in-house, which combines the fixed output of a single vaporizer (3% oxygen) in variable ratios with pure oxygen so that the isoflurane concentration for each rat was individually adjustable. Each rat was injected with 1 mCi11C-PK11195 through the caudal vein. Each rat was scanned for 30 min with its brain centred in the axial and transaxial fields of view (Explore VISTA Micro-PET/CT scanner from GE Healthcare, USA). The image data were sorted into three-dimensional sinograms, followed by attenuation and scatter corrections, Fourier rebinding and two-dimensional ordered-subset expectation maximisation reconstruction with two iterations and 50 subsets. The image pixel size was 0.385×0.385×0.385 mm. For semiquantitative evaluation, the maximum uptake values were automatically measured three times in the ROIs (hippocampus and prefrontal cortex) and the uptake ratio was calculated.19
Immunohistochemistry
Rats were deeply anaesthetised and sacrificed by cervical dislocation. Brains were removed, fixed in 4% paraformaldehyde for 4–6 hours, dehydrated with 30% sucrose in phosphate-buffered saline (PBS) overnight, embedded with optimum cutting temperature compound, cut into 10 μm thick sections serially using a freezing microtome (CM1950, Leica, Germany), mounted onto superfrost plus glass slides and stored at −70°C for immunohistochemistry. Six sets of serial sections were obtained from each brain, and one of them was used for immunostaining.
Immunohistochemistry was performed as previously described.20 After being washed in PBS, endogenous peroxides were blocked by incubation in 0.3% hydrogen peroxide in 50% methanol in distilled water for 30 min. Sections were washed in PBS and preincubated in a blocking solution containing 5% bovine serum albumin for 15 min at room temperature. Then sections were incubated in a humidified chamber with the rabbit primary antibody against CD11b (1:100; Boster Biological Engineering, China) in PBS overnight at 4°C. After several washes in PBS, sections were incubated with biotinylated goat anti-rabbit secondary antibody (1:100; Boster Biological Engineering) for 60 min at room temperature, followed by incubation with the streptavidin-biotin-peroxidase complex method (Boster Biological Engineering). Following the PBS rinsing, the antigen-antibody complexes were visualised using diaminobenzidine (Boster Biological Engineering) as the chromogen. The immunohistochemical controls were performed as above, but with the omission of the primary antibodies. Eight sections were selected randomly from the hippocampus and prefrontal cortex in both groups. Six images were taken from each section using ×10 magnification randomly. Images were captured using a microscope (DM1000, Leica) and the DM2000 Image Acquisition and Analysis System. Image-Pro Plus software (V.6.0, Media Cybernetics, Silver Spring, MD, USA) was used for statistical analysis. Optical density (OD) value was used to quantify the CD11b positive cytoplasm. The OD values were positively correlated with the strength degree of the positive expressed area. OD value=integrated optical density/positive results area.
Statistical analysis
All analyses were performed using the Statistical Package for the Social Sciences (SPSS V.20.0). Quantitative variables in group comparisons were analysed using the independent t-test. Sex differences were analysed using the independent Χ2 test. Pearson correlation was used to examine the relationships among behavioural, immunohistochemistry and micro-PET/CT measures. Because there was no significant sex difference in any of the preliminary analyses performed, the male and female subjects were collapsed to enhance statistical power. Statistical significance was set at p<0.05 (two tailed).
Results
Behavioural measures
As shown in figure 1A, the treatment group had a significantly higher locomotor activity than the control group (t=5.239,p=0.002).
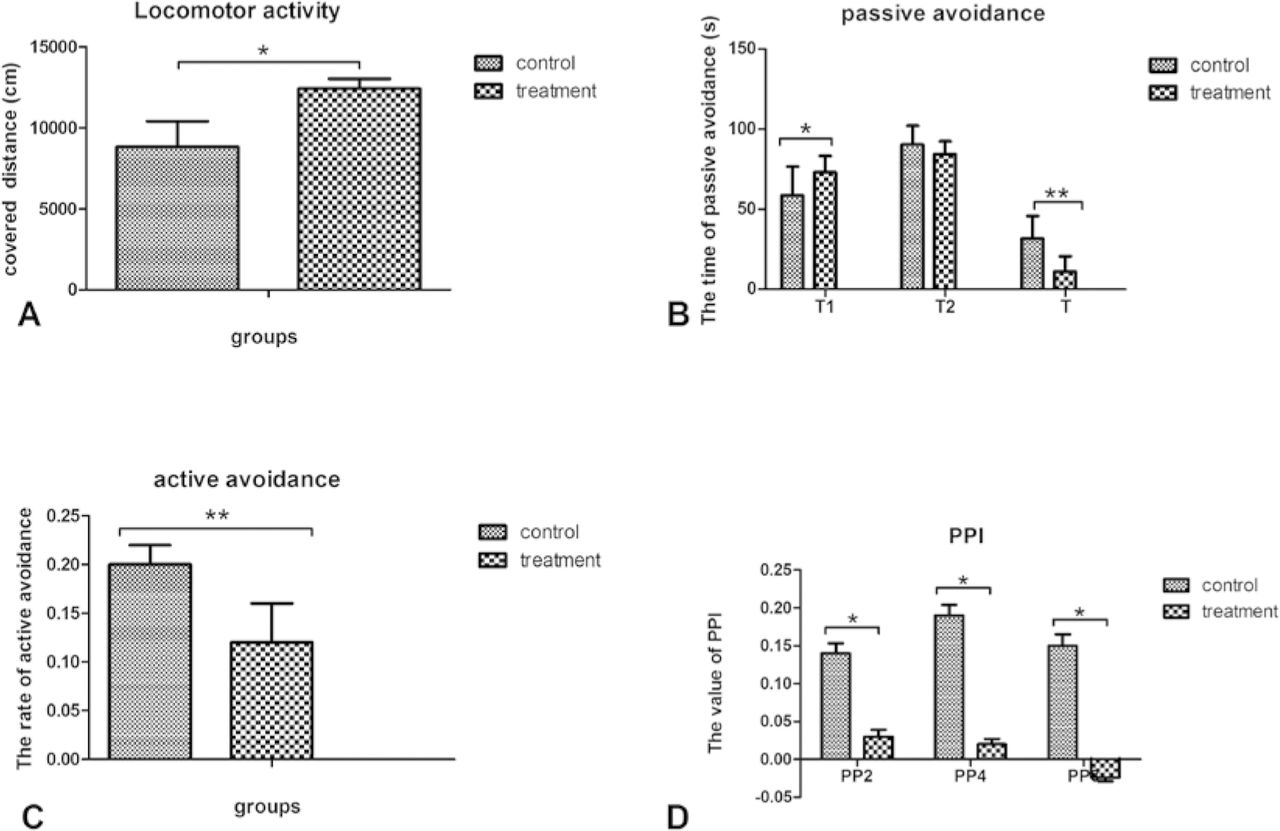
Group comparison of behavioural measures. (A) Locomotor activity as measured by the distance travelled in the open field during 60 min of treatment. (B) Latency in the passive avoidance task. (C) The rate of active avoidance conditioned response. (D) Prepulse inhibition (PPI)deficits.
During the passive avoidance task, the treatment group showed a significantly longer latency on the first day (T1) than the control group (t=2.548, p=0.036). However, there was no difference in latency on the second day (t=−1.452, p=0.161) between the two groups. Consequently, the treatment group showed significantly lower T values than the control group, reflecting their learning and memory abilities (t=−2.618, p=0.038) (figure 1B).
During the active avoidance task, the treatment group showed a significantly slower rate of conditioning than the control group (t=−4.749, p=0.005) (figure 1C).
For the PPI test, the treatment group showed significantly lower PPI values than the control group across different prepulse intensities (72dB: t=−2.603, p= 0.028; 74 dB: t=−2.520, p=0.047; 78 dB: t=−7.718, p< 0.001)(figure 1D).
Micro-PET/CT scan
The isoquinoline PK11195 binds to the translocator protein (18 kDa), the expression of which is believed to be enhanced in microglia during activation.21 11C-PK11195 uptake ratio was significantly slower in both the hippocampus (t= −4.462, p= 0.001) and prefrontal cortex (t= −3.990, p= 0.003) in the treatment group compared with the control group (figures 2 and 3).
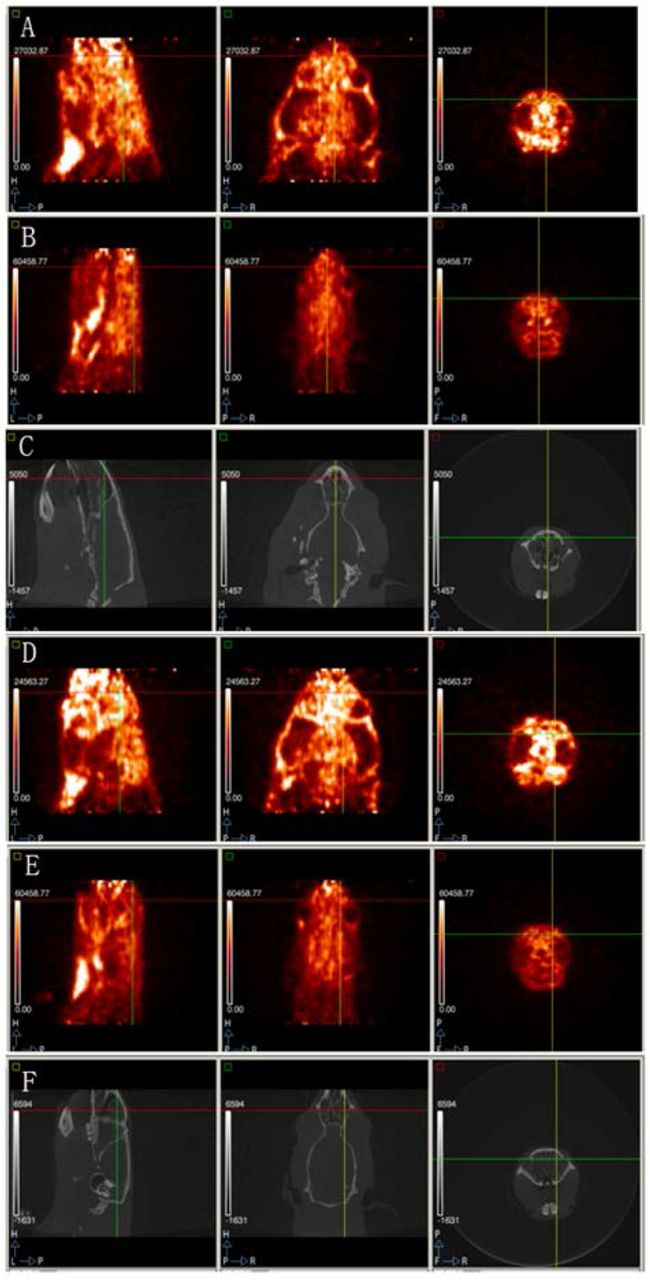
(A) the prefrontal cortex PET scan of the treatment group; (B) the prefrontal cortex PET scan of the control group; (C) the prefrontal cortex CT scan of the treatment group; (D) hippocampus PET scan of the treatment group; (E) hippocampus PET scan of the control group; (F) hippocampus CT scan of the control group. Each panel shows the sagittal, coronal and horizontal positions from left to right. Brighter areas reveal a higher uptake ratio in the brain.The treatment group combined more radioactive material than the control group in both brain areas.

A:11C-PK11195 uptake ratio in the prefrontal cortex B:11C-PK11195 uptake ratio in hippocampus note: p1-p5 means 5 time points11C-PK11195 uptake ratio in the prefrontal cortex; h1-h5 means 5 time points11C-PK11195 uptake ratio in hippocampus.
Immunohistochemistry
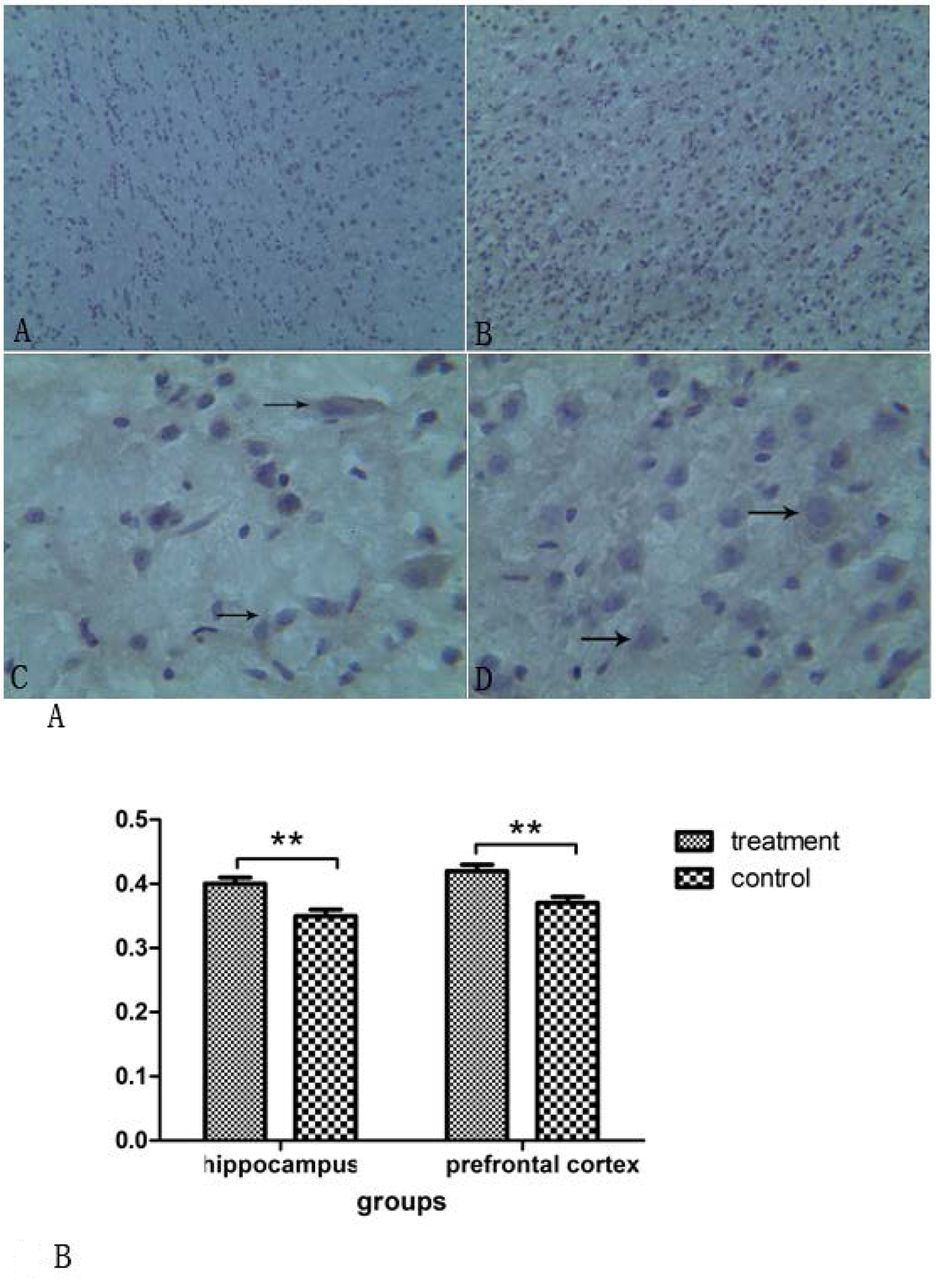
Most of the microglia cells in the control group were in a resting state. The PET scanning showed round or oval cell bodies(figure 4A: A-C). In the treatment group, most of the microglia cells were activated (figure 4A: B,D). There was a significant difference in the OD values of the two groups in the hippocampus and prefrontal cortex areas (hippocampus: t=8.204, p< 0.001; prefrontal: t= 6.995,p< 0.001)(figure 4B).

(A) CD11b-immunopositive cells in the control group (A, C) and thetreatment group (B, D) Group comparison of OD values in the prefrontal cortex and hippocampus
Correlations data
Table 1 shows the correlations among the behavioural tests, micro-PET/CT scan and immunohistochemistry. There were significant positive correlations between the decay rate of the prefrontal cortex and PP2, PP4, PP8 in behavioural tests. There were significant negative correlations between the immunohistochemistry OD value of the prefrontal cortex brain areas and PP2, PP4, PP8 in the behavioural tests. Locomotor activity was significantly negatively correlated with the decay rate of PET/CT, and positively correlated with the OD value. Passive avoidance was significantly correlated with hippocampus area. Active avoidance was significantly negatively correlated with the OD value and positively correlated with decay rate of PET/CT (table 2). Table 3 showed a correlation between the micro-PET/CT scan and immunohistochemistry. There were also significantly positive correlations between the OD value and decay rate of PET/CT in the same brain area (table 3).
Correlations analysis between behavioural measures and PET/CT11C-PK11195 rates of decay, as well as immunohistochemistry OD values
Correlations analysis between PET11C-PK11195 rates of decay and immunohistochemistry OD values
Discussion
Main findings
This study aims to investigate microglia activation in the offspring of rats exposed to Poly I:C during pregnancy using 11C-PK11195 micro-PET/CT and immunohistochemistry. Our study demonstrated activated microglia in the prefrontal cortex and hippocampus areas, and the microglia activation was associated with SCZ-like behaviours.
We choose 56 days’ offspring because: (1) the age of 56 days means a mature state of SD rats; and (2) the experimental rats in the literature are 56 days.7 In the preliminary experiment, we tried to use Iba-1 and CD11b, respectively, as the primary antibody for immunohistochemistry. The result was that CD11b was more suitable for this study.
As shown, the branches and processes of the microglia cells derived from the Poly I:C exposed SD rats were different from those of the control group. While microglia cells from the control group rats showed a more highly branched arborisation, indicative of a non-inflammatory state of microglia, the offspring from the Poly I:C-treated rats were not characterised by none or fewer branched cells, as characteristic for an activated and inflammatory or phagocytic state of microglia.22 23Juckel and colleagues showed that offspring microglia from Poly I:C-treated mothers were morphologically characterised by a reduced arborisation indicative for a status of higher activation compared with the offspring microglia from vehicle-treated mice.23 A reduction in ramified microglia and the overactivation of microglia cells at a young age may have a detrimental impact on the response by these animals to future neurological insults or injury. Williamson 24and colleagues suggest that adult microglia arise exclusively from the population of resident macrophages, meaning a reduction in ramified or resting microglia from an early age may have an enduring impact on the function of these resistant macrophage-like cells.
In addition, there were significant positive relationships between the results of the micro-PET/CT scan and those of the immunohistochemistry. Our findings suggest that prenatal Poly I:C-induced SCZ treatment showed active microglia in the hippocampus and prefrontal cortex brain areas. In a study of descendants of mice exposed to Poly I:C at embryonic day 9, a higher number and activation of microglia cells in the hippocampus, but not in the prefrontal cortex, existed at postnatal day 30.25 Several previous studies with C57BL/6J mice reported that active microglia cells were present in the prefrontal regions in chronic stress, and chronically available alcohol.26 The difference in the brain areas of the active microglia might be explained by the differences in the species of mice. Increasing evidence suggests a role for microglia in normal synaptic plasticity mechanisms within the adult brain, including dendritic spine retreatment and elimination.27 Some researchers have demonstrated that microglia primed as a consequence of neonatal infection and that this priming leads to aberrant production of IL-1 specifically during learning in adulthood.24 ‘Priming’ of microglia, as in a fetal inflammatory challenge such as that provided by Poly I:C, may result in exaggerated response to a subsequent insult,28 due to increased production of cytokines compared with the ‘unprimed’ microglia.29 It is thought that this overproduction of cytokines due to a subsequent and undetected insult or infection could be the underlying mechanism leading to postnatal cognitive abnormalities related to the prefrontal cortex.30 Importantly, the resident microglia of the hippocampus are a necessary component of normal memory formation.24Vinet and colleagues indicated that ramified microglia screen their microenvironment and additionally protect hippocampus neurons under pathological conditions.31
Our study found that the activation of the microglia was associated with SCZ-like behaviours. These findings are consistent with several previous findings.32–34Ito and colleagues reported that changes in CA1 hippocampal c-FOS expression (an immediate early gene measure of neuronal activity) and in excitatory postsynaptic currents (measured in slices maintained in vitro) led them to conclude that the altered processing of object-place recognition was of hippocampal origin. Richetto 34and colleagues found that prenatal Poly I:C-induced immune activation led to an age-dependent decrease in GAD65 and GAD67 expression in the prefrontal cortex and hippocampus, two brain areas critically involved in (spatial) working memory.35 36 In a study of 16 patients with SCZ, functional connectivity indices in the left dorsolateral prefrontal cortex and the right hippocampal cortex were positively correlated with accuracy during the working memory task, while the connectivity strength in the right dorsolateral prefrontal cortex was negatively correlated with measures of the severity of the working memory task.37 Furthermore, a recent study38 found that high-risk individuals showed decreased activation compared with healthy controls in the inferior frontal and prefrontal cortex during a semantic working memory task.
In postmortem studies, activation of microglia has indeed been found in several patients with SCZ, especially in the frontal regions of brain.39–42 A previous study of 10 patients with recent-onset SCZ reported that compared with control subjects, the R-11C-PK11195 binding of total grey matter was increased in patients with SCZ.43 It is possible that microglia activation may play a possible or potential role in the neuropathology of SCZ. Recently, two PET studies on SCZ versus controls showed lack of significant difference in neuroinflammation in treated patients with SCZ in the midst of a psychotic episode. The neuroinflammatory processes may take place early in disease progression or are affected by antipsychotic treatment.44 45
Compared with previous studies, the strengths of this study included the prenatal Poly I:C-induced immune activation treatment, the use of 11C-PK11195 as a trace of micro-PET/CT scan, combined with CD11b immunohistochemistry and behavioural assessment.
Limitations
However, the study also has several limitations: (1) it was only studied at one time point at day 56, so it is not possible to conclude how long this activation of microglia might exist; (2) a causal relationship between microglia and cytokines cannot be drawn from the study design.
Implications
Future studies are needed to dynamically track the in vivo microglia activation from an embryonic state to adulthood in the Poly I:C prenatal infection animal model, examine the levels of cytokines in the cerebral spinal fluid and peripheral tissue and investigate the intervention strategies to prevent and treat SCZ-like behaviours.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Flowchart of the study.
Supplemental material
References
Footnotes
XL and XT contributed equally.
Contributors XL and XT were responsible for the analysis and interpretation of data for this paper. All authors contributed to the writing of the paper.
Funding Funding for this study was provided by the National Natural Science Foundation of China (No 81571318 to XQS; No 81371472 to LXL; No 81401110 to XL), the Science and Technology Planning Project of Health and Family Planning Commission (No 201501015 to XQS), the International Science and Technology Cooperation Program of Henan (No 162102410061 to XQS), the Henan Province Union Fund Project (162300410275), the Zhengzhou University doctor team project, and the Youth Fund of the First Affiliated Hospital of Zhengzhou University (to XL and LJP).
Patient consent Not required.
Ethics approval The protocol was approved by the Ethical Committee of The First Affiliated Hospital of Zhengzhou University.