Article Text

Statistics from Altmetric.com
Introduction
Although the content of one’s consciousness changes through time (eg, personality changes), their identity or psychological continuity still, somehow, remains the same. In fact, typical human self-continuity—the unity of consciousness over time—spreads out over a period of hours, from one episode of dreamless sleep to a waking state. I wake up, eat breakfast, take a shower, get dressed, go to the institute and start the routine work of my scientific life. I am conscious the whole time, and the experiences I have at each phase of my morning, belong to a single stream of consciousness.
However, this smooth subjective occurrence of self does not function for some people. In some patients, like those with schizophrenia, self-continuity abruptly ceases. They experience a kind of divorce from reality with various behavioural disturbances such as hallucinations, delusions and catatonia. In practice, the patient experiences a weird sense of losing their identity. It seems that they are unable to preserve their identify of self with the outside world. In other words, they lose contact with the reality of the world.
There is also a large body of literature suggesting that those with schizophrenia turn away from the outside world and become concerned with their own subjective state.1 By a closer look at some of the thought disorders of schizophrenia, we notice that a metaphoric glue in the stream of consciousness is missing. This includes derailment or tangentiality or knight’s move thinking—a failure to follow the social norms of conversation and not sticking to the overall theme of the discussion, or pressure of speech— a speeding up of the flow of speech. Besides, there is evidence indicating that their perception of the world differs qualitatively from that of a normal individual2; this, in consequence, leads to a deprivation in their gratitude towards the outside world.
This is exactly the point which we should notice more. Why do they lose contact with the outside world? What happens inside the minds of these individuals that makes them disconnected? Is it just neurodegeneration in a region of the brain (such as in Alzheimer’s) or lack of a special kind of neurotransmitter (such as in Parkinson’s disease)? Or, is it only a genetic problem? Although many research institutions that investigate the aetiology of schizophrenia have concentrated on organic deficits, they are unable to reach a consensus on why these people lose their thread of self.
Before answering these questions, one of the main locomotion theories after the dawn of the cognitive revolution in the mid-20th century—the reafference principle (von Holst and Mittelstaedt)—should be reviewed. One may presume that the answer to these questions lies within a better understanding of this theory.
Von Holst and Mittelstaedt’s principle of reafference
Having been inspired by Johann Georg Steinbuch3 and von Helmholtz’s ‘Willensanstrengung’/‘effort of will’ concept,4 von Holst & Mittelstaedt5 were the first to suggest in a thorough manner, and with experimental evidence, that motor-to-sensory feedback has a leading role in regulating animal behaviour and actions (in contrast to James’ reflex theory,6 which assumed that all incoming signals must pass through a sensory medium, and that information which flows within the brain are guided in the sensory-to-motor, rather than in the motor-to-sensory direction).
This groundbreaking theory was published as a profound break from behaviourism which described behaviour as a set or sequence of stimulus–response actions (for instance, the reflex arc concept, which was generally accepted by scholars at that time). Moreover, it seems that the main incentive upon the advent of this theory was a response to early 20th century scraping and locomotion works by Sherrington and Graham Brown in vertebrates.7 8 Sherrington determined that a scratch reflex occurs with mechanical stimulation (eg, skin tickling) in a wide saddle-shaped region of the upper part of the body by transecting the cervical spinal cord of the dog. In other words, he concluded that no internal feedback implementing an action is needed. These discoveries by Charles Sherrington brought severe criticism to von Holst and Mittelstaedt’s hypothesis; he attacked this idea and concluded that it ‘remains unproven’.9
It appears that Sherrington did not view afferent signals as complementary rather than exclusive mechanisms for organisms’ acts. It is worth noting that this way of thinking was not restricted to Sherrington or Brown. Most scientists who strive to explain human behaviour, make the mistake of assuming that the brain is enough to model it. They ignore the environmental variables which interact with the brain in a complex multifactor cosmology to create the behaviour of an organism. However, it is through these interactions that we perceive the world. The author supposes that these kinds of models which were proposed compensated for the lack. Here is a brief revision of this theory.
The reafference principle
Von Holst and Mittelstaedt first proposed this model in 1950. This model attempts to address the question of how an organism’s inner and outer world can be perceptually stable, despite the fact that the sense organs are activated as we walk around in the world. They believe that our brain always makes a copy (they call it the ‘efference copy’) as a default picture and it always compares that copy to the outside world. The efference copy, as they believe, causes a strictly correlated neuronal process which is stored in the adjacent ganglia after a certain time delay. The flurry of pulses that flood through the periphery stimulates its effector, which contributes to afference. In essence, this interacts with the efference copy (in anticipation for). There is a close relation between the central nervous system (CNS) and the environment. They conducted animal experiments to demonstrate this theory. There is, therefore, an ideal internal ‘model’ which reflects the real state of sensory data, according to the principle of reafference. When a goal is set, the brain arrays an inner model and the sensor system must play a kind of game to change the external data before it is formed to match the internal model and sensory data.
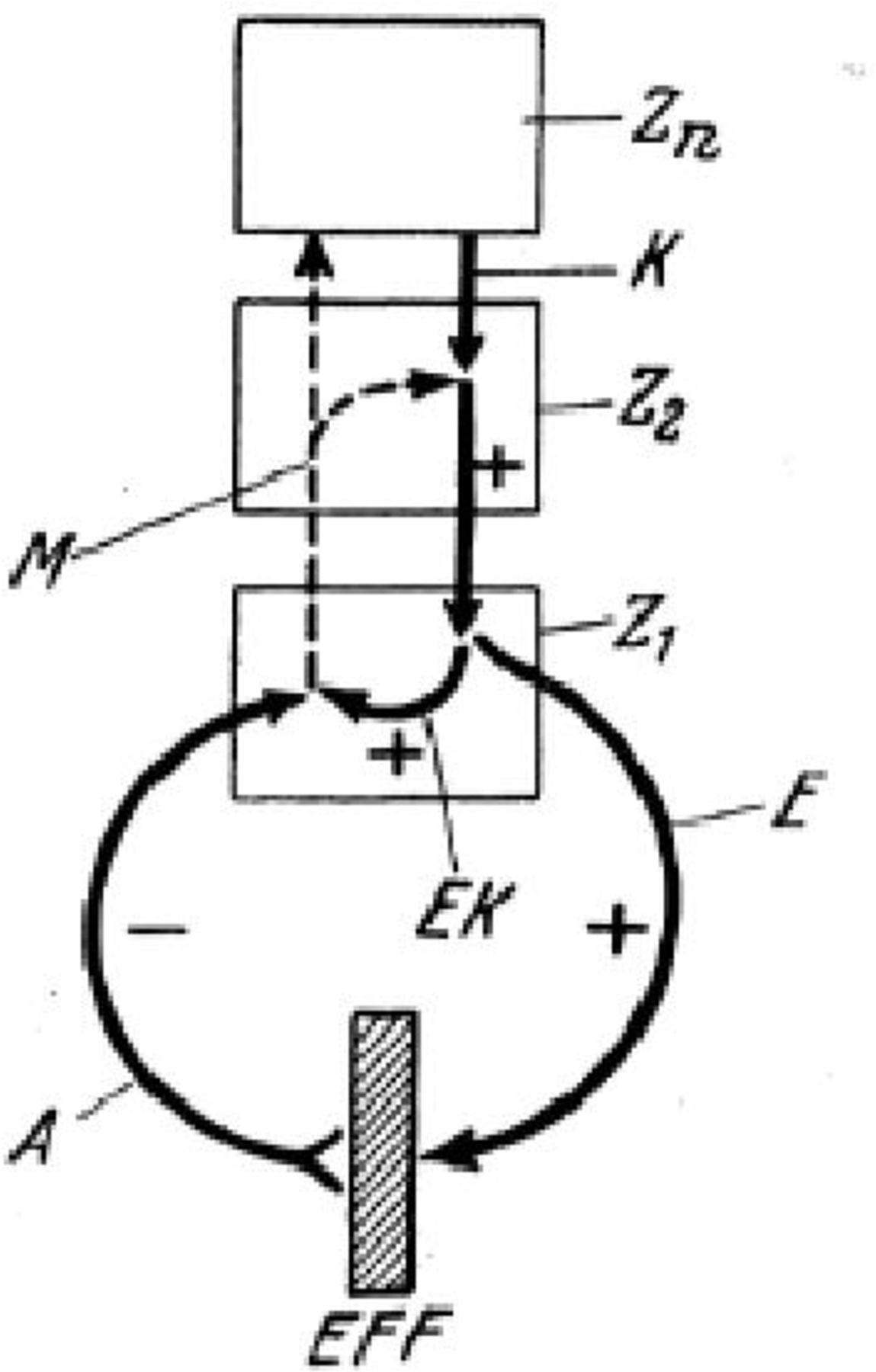
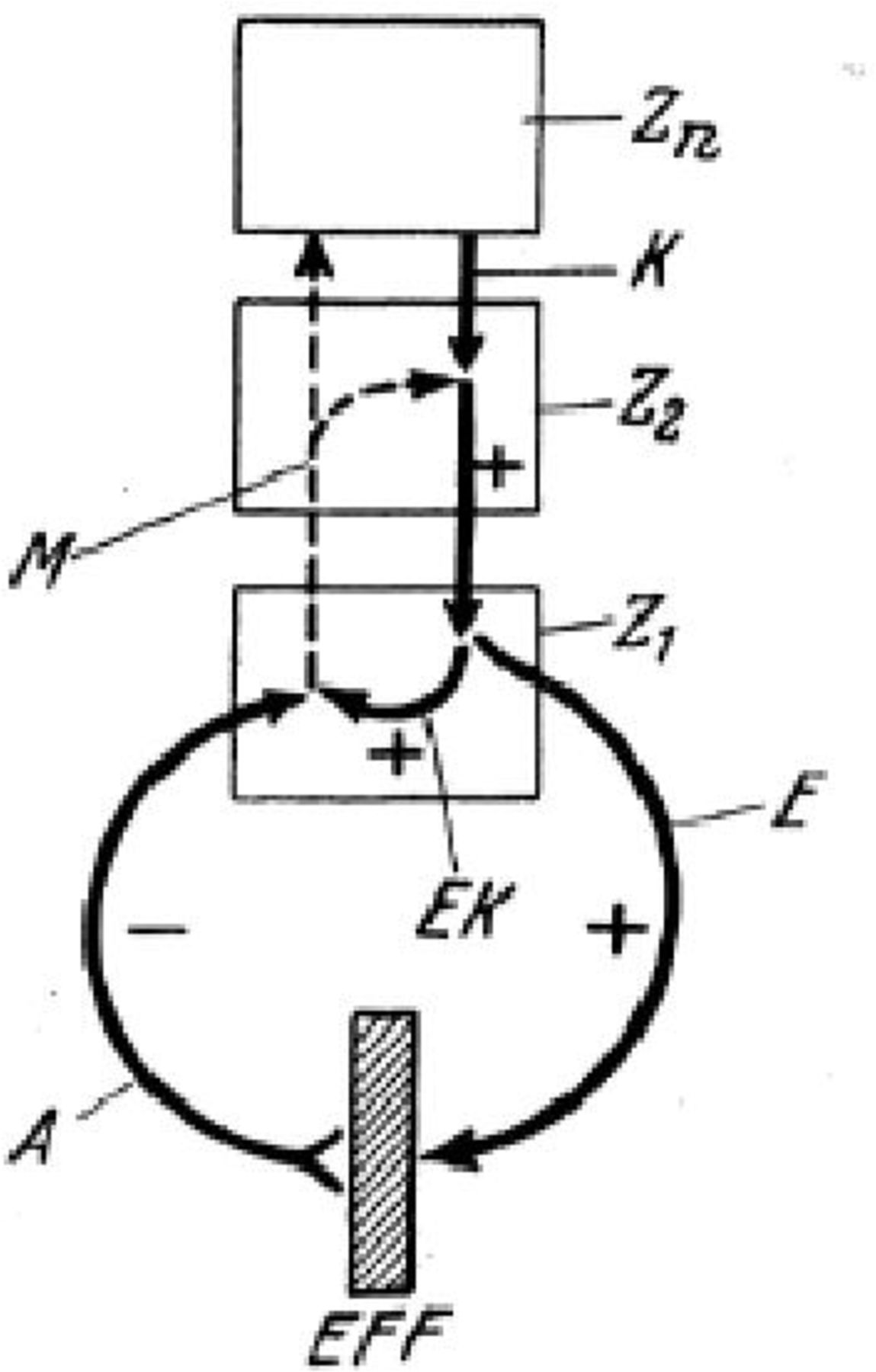
As this model is refined, that is, the brain continues to adapt until there is a reduction of the discrepancy between the inner model and the sensory data and the objective (figure 1).
{kind=link}
General schema for the principle of reafference. Borrowed form von Holst & Mittelstaedt (1950). A command K is sent from a higher centre of Zn to the lower centre; a neural command from there is put at the executive centre (Z1). This command in turn interfaces an efference E to an effective centre (EFF). The efferent instruction E (efference copy (EK)) has been stored at the same time. After movement, the afferent signal A (or 'reafference') is sent to Z1, where the EK is compared with this reafferent signal. A (with a minus sign) similar to EK (with a plus sign) cancels all signals, and the sensorimotor loop is done. If EK and A do not correspond, Z2 and Zn are sent a message (M) for correction or reinitiation of the sensorimotor cycles before it ends.
From von Holst and Mittelstaedt’s viewpoint, each organism’s CNS behaves in an automatic manner—either directly or indirectly—with the surrounding environment in the world. For these authors, there is a lawful relationship which should be discovered. At the beginning of their notorious article—The Principle of Reafference: Interactions Between the Central Nervous System and the Peripheral Organs—while introducing the principle of reafference, they try to draw our attention to the importance of this concept:
A major question for the physiology of the CNS has always been this: what lawful relationships hold between impulses which are generated by external stimulation and travel inward into the CNS and those which—either directly or indirectly—reemerge from it; that is, the question of the relations between afference and efference? The CNS has been characterized as a sort of automat, which reflex delivers a given ticket when a particular coin is inserted in it. For simple protective reflexes—like sneezing and withdrawal from painful stimuli, this idea is easy to accept; for more complicated reactions such as balancing and orienting responses, the same conceptual scheme has been advocated. Even rhythmic locomotor patterns can be understood in these terms if one assumes that each single movement reflex evokes its counterpart and that each component sets off its successor in time (reflex chain theory). The higher forms of behavior, which are modifiable through experience, are subsumed under the same rubric by way of “conditioned” reflexes.
However, this hypothesis seems wrong to many behaviourists. For them, it seems unscientific that the CNS is an active organism with potential activities without external stimuli. In this case, for example, if we take sleep and rest as special modes of CNS activity, it is hard to limit anything to the notion that ‘causes’ of some key occurrence remain ‘stimulation’. But von Holst and Mittelstaedt think that this hypothesis could be dismissed because of our limited thoughts of resistance and laziness. They encourage us to see the situation from a completely different angle.
Von Holst and Mittelstaedt began by asking the following question: ‘How can organisms respond differently to exafferent and reafferent stimuli, even when these are quantitatively and qualitatively alike and when they are superimposed in the same channel? For instance, how do animals overcome stabilising reflexes during voluntary actions? Take, for example, the stability of space by our vision. If you start moving your eyes passively up and down, objects also become unstable and you feel they are moving. Furthermore, patients who suffered from a brain injury are no longer capable of moving their eyes. They experience spatial instability in their visual field. Or, hand–eye coordination in young children—the ability to use their muscles and vision in tandem—necessitates the development of visual skills, like visual acuity and muscle skills. When the two work together in a coordinated way, young children begin to increase their ability to pick up, hold and manipulate objects. For experiencing stable vision, voluntary impulses and visual signals have to be coordinated.
It should be noted that their theory is not restricted to an organism’s movement. They bring another illustration in performing the principle of reafference which can be traced to visual processing and its relation to memory. The traditional notion of visual processing affirms that when we identify an input (afference) as something, we compare our perception of a new stimulus with items stored in episodic memory. It is analogous to finding a book in the library. However, this system does not function for some who suffer from visual agnosia, a condition in which patients are unable to recognise the class to which an object belongs. These patients have difficulty recognising a variety of visually presented objects. They cannot name them or group them into semantic categories. Remarkably, these patients can copy drawing of objects and they can match the pairs of drawing as the same or different (however it is very slow in comparison with healthy subjects) even using objects they do not recognise. It seems these patients have normal visual input processing but the stored memories are hypothesised to be damaged.
As the closing comments in this section, it should be maintained that the reafference principle can be applied throughout the CNS from the lowest mechanisms such as internal and external control of the limbs, relations of diverse parts of the body to each other, to the highest like orientation in space, perception, illusions and many others. Furthermore, one may suppose that it can bond ‘low-level machinery’ nerve physiology, to the ‘high-level system’ in behavioural science.
The lack of a timing system
Having scrutinised the aforementioned fundamental theory (the principle of reafference), one may realise that there should be another system which functions in this theory. The model talks about the interaction of the brain with the outside world, but there is no mention of a timing system. In other words, if there is an anticipatory system in the brain which always compares the outside world (aim) with the efference copy, how long does this temporal interval take?
Precise timing of actions is the cornerstone of our movements. Time determines whether our actions are completed or not. Therefore, the brain is trying to solve a timing challenge—the timing of fine body actions (motor timing).
Have you considered how a dancer achieves accurate motor timing under various conditions? We take for granted how humans synchronise their movements with ease to nearby moving objects. This requires a highly timed processing of the perception–action arrangements underpinning movement control. Motor activities must be accurately timed for their proper execution; completing one component too early or too late could make the action ineffective. Temporal information in the scope of milliseconds is vital for a wide range of cognitive and motor processes. Time perception is evolutionary adaptive machinery that facilitates the ability to predict and anticipate events at an appropriate time, as well as organise and plan sequences of actions for fight or flight.
Distinguishing the temporal connection between visual stimuli, for example, is a determinant for the direction and speed of moving objects. Likewise, the capability to accurately perceive and generate temporal structures of sounds is essential for distinguishing and producing different languages.
According to many studies, perceiving appropriate motor timing and precise timing is vital for the preparation and execution of most motor responses.10 These two play an important role for humans to perform optimally in the environment. For instance, one has to foresee when sensory events will occur and estimate how long these events will endure to move their body, cooperate with others and act towards environmental stimuli. However, sometimes this motor timing mechanism in which organisms plan the future with precise timing and foresee the results of actions does not function efficiently. Behavioural and neuropsychological data indicate that hyperactive children and patients with Parkinson’s and Huntington’s disease11 12 show poor intertemporal competence (ie, poor ability to associate events, which are separated by time, paced and self-paced finger tapping, and simultaneous and sequential acts).
Moreover, in prehensile movements, proper coordination and precise motor timing of the arm, hand and trunk are required. For example, when we want to grasp something, the time from movement initiation to first object contact and time taken from first contact with the object until the object is touched should be considered. However, the precise motor timing of these acts is taken for granted in our speedy life.
One of the reasons that visual and motor structures evolved together is to support life’s acts. Even in an oculomotor system—the system of eye movements—timing is crucial. For instance, in one type of voluntary eye movement in primates, saccadic eye movements (reading this sentence, for example), the eye position signal initiates to change at least 100 ms before the onset of a saccade and does not reach a veridical value until about 50 ms after a saccade finalises.13 14
It is believed that the cerebellum plays a leading role in the timing of acts. This idea originates from the fact that the cerebellum has several connections with important cortical and subcortical regions of the brain such as the primary motor cortex (M1), the supplementary motor area, the cingulate cortex and the basal ganglia. Furthermore, it has been shown that cerebellar lesions lead to behavioural problems in a range of timing tasks.15
In describing the importance of precise motor timing, we can observe professional athletes, who build their careers on motor timing mastery and hone their proficiency to the very edge of human capability. For example, a sprinter must foresee the firing of the starter’s gun to get a jump on the competition. How does an individual decode such temporal aims into an action? A wealth of literature on motor timing suggests that temporal control arises, at least in some situations, directly from motor control circuits in the brain.16 17 Interestingly, many of our tasks have temporal aims or require precise temporal information for their successful accomplishment.
It is worth noting that motor timing is not restricted to just acts in the outside world. Each organism on the basis of its anatomical structures, such as the jaw or tongue, has a predictable relationship in the relative timing of their movements. Timing in prediction and anticipation should take the form of direct motor behaviours, such as anticipatory licking or blinking. Even with internal activities, controlling movements fundamentally encompasses activating motor units at the right time. As a matter of fact, a motor learning mechanism without considering temporal specificity would be a poor one. For example, a bird to pick up a piece of seed must open its beak and close it at exactly the moment when the beak surrounds the target. We notice that it is an action that must be precisely timed if it is to be a successful act.18 19
As another example, in language impairment studies, evidence indicates that fine-grained temporal integration in a language system is fundamental to speech and language.20–23 The impairment in certain aspects of speech and language such as stuttering could be due to the impairment in precise vocal timing and perhaps, also in intonation abilities among these persons.
Our most detailed neurobiology of time in sensorimotor function comes from experiments in animal models. These experiments have shown how neural activity in various cortical and subcortical brain regions underwrites the overall ability of an animal to produce timed behaviours.24 25
Brain regions underlying cognitive processes such as attention, memory and decision processes work together with the neural timer as part of a network serving accurate timing of movements. According to some studies,26 two brain regions are involved in this network—the cerebellum and prefrontal cortex. Ivry15 hypothesised that the cerebellum functions as an internal clock that serves to time precise temporal associations between events in both motor and perceptual domains. For instance, in dysmetria (a kind of ataxia which is a neurological deficit), there is a lack of movement coordination characterised by the undershoot or overshoot of an intended position. This is due to dysfunction of the cerebellum. Another study reported that patients with impaired cerebellum are unable to judge the accurate duration of an auditory stimulus.27
Likewise, the supplementary motor area has generally been assumed to play a prominent role in the coordination of the homologous limbs. However, this cannot be the whole story because the coordination of limbs needs accurate timing. It is believed that the cerebellum is generally responsible for motor timing synchronisation—integrating motor commands and sensory information to help coordinate movements.28 29
In recent years, imaging studies have linked a large number of brain areas with motor timing, including the basal ganglia (insights coming from studies on Parkinson’s disease), the cerebellum (especially Purkinje cells as inhibitory projection neurons) and several prefrontal cortical structures.30 31 However, some researchers believe that it is not appropriate to restrict motor timing to a specific region. They have proposed the cortical circuits which function in motor timing. They argue that the temporal control of behaviour likely spans the whole spectrum from the sensory cortex, to higher-order associative areas, to the motor cortex. For example, information about the predictable time of sensory events such as visual signals has been detected in the primary visual cortex of monkeys and rats.32–35 These findings suggest that the primary visual cortex may have a leading role in the construction of visually cued timed actions.
Another criticism, which has been levelled at motor timing research, is that many of these experiments are very similar to finger-tapping tasks. It seems that for the researchers in this field, a hammering motion of a finger and an up-and-down motion of the jaw are very similar. However, some believe that persons who are stable timers in a tapping task are not necessarily stable timers when they perform a drawing task. In other words, there is no meaningful correlation between tapping and drawing in terms of timing precision. This suggests that timing behaviour is exclusive to the nature of the task, and thus weakens the idea that timing is a generalised capability that can be executed on diverse types of tasks.36 37
Having highlighted the vitality of motor timing in an organism’s survival, many other questions still arise: how does the brain know the right timing for an action? How does the brain perform such relatively accurate timing with which human and non-humans are able to survive in the complexities of the outside world?
Elucidating schizophrenia
One may presume that the answer to this enigmatic mental illness called schizophrenia lies in an under-represented concept among experts in this field: time. As it mentioned in the previous section and I would like to generalise it to those with schizophrenia, one of the causes of schizophrenia and depression is due to an inefficiency in the precise timing system of the brain. By this, I mean there occurs a deficiency in temporal integration between the comparator and anticipatory mechanisms in the CNS. If this precise timing system (or temporal integration) becomes affected, the abilities to immediately experience and consciously perceive the world both get disrupted. Put simply, an altered inner time consciousness gives rise to an altered self-consciousness, with this alteration being most noticeable in patients suffering from these disorders. For instance, based on personal reports of these patients, there exists an altered time and self-perception as well as an incapability to deal with daily life. Some patients report their daily states as feeling a ‘heaviness’, whereas the passing time is perceived as kind of ‘gluey’. The person with depression gets stuck in circling thoughts which join him to the past. These recurrent thoughts become significant and lead to a feeling of inertia as well as to a ‘disconnection’ from the present world.
Reduced motivation is another typical symptom in depressive patients in which an altered subjective temporality is experienced. For them, the future becomes unachievable. An orientation towards goals in the future is impossible. This often leads to a feeling of guilt. It seems that creating a new self is dreadful. The person concerned becomes a ‘captive of the past’ within a decaying now. The transient time of the ‘outside world’ is seen as being detached from the subjective time of the perceiving self.
Even in some symptoms of schizophrenia like auditory hallucinations (patients may experience one or more fantasised voices, which may comment on their activities, or talk directly to them), the inefficiency in the precise timing system of the brain between the comparator and anticipatory mechanism can be tracked. But, what are the mechanisms involved in auditory hallucinations? The normal experience of ‘inner speech’ offers a clue to this phenomenon. The capability to control one’s behaviour by means of self-directed speech progresses in early childhood, when children first talk out loudly to themselves before learning to internalise this process. This situation reaches a climax in adulthood in the capacity for mature, verbal thought. Even in adulthood, this kind of contemplation is accompanied by ‘subvocalisation’. Subvocalisation consists of a covert activation of the speech muscles that can be identified by electromyography. It has been notorious for many years that auditory hallucinations are also accompanied by subvocalisation.38 39 This observation has inevitably suggested to many researchers that auditory hallucinations occur when inner speech is mismatched to an external source .40 41 Many of these symptoms are seen in schizophrenia with the same impact on the victim. In most of these cases, there is a kind of misidentification, an alienated self-consciousness. In alienated self-consciousness, individuals experience their own conscious events as those of another agent and consequently fail to recognise their own thoughts or feelings as their own.
Here is exactly what should be highlighted and what most of these theorists have not considered: could it be that vocal hallucination is just a kind of subvocalisation in the patient caused by a temporal integration problem? Could we say that the patient hears his own voice, but due to a temporal integration problem between the comparator and anticipatory mechanism of the brain, he feels that the voice is coming from somewhere else?
For example, consider this: auditory hallucinators usually do not designate voices as qualitatively or phenomenally akin to ordinary auditory or speech perception. Quite often they state that their experience of the voice is not dissimilar with their normal experience of their own inner speech. So, what leads them to judge that their experience of voices is a representation of another’s speech? According to the hypothesis mentioned in the previous section, this would be due to the ‘verbal imagery’ of these individuals (ie, all of us have a copy of our voice in our head/efference copy). However, this seems strange due to dysfunction in the precise timing of the brain between the main copy/efference copy (the verbal imagery) and the new ideas which cannot be matched (in the comparator system) with the centre of identity (inclusive image). Therefore, the patient experiences it as an alien source.
Individuals with schizophrenia lose the capability of preserving the self and perceiving the present time becomes a crucial phase of their disorder. In fact, obliviousness towards time paralyses the self in these patients, affecting them to feel hollow and unable to function in the present. For instance, those with schizophrenia occasionally think they are children while they are grown-ups, or otherwise forget themselves in past time. It looks like that they are forcibly cut off from their present existence and experiencing a loss of identity. However, disconnection of identity with the present time is not restricted to mental illness. Soldiers who come back from war often report confusion as who they really are as well. This problem happens because the nature of the self is eager to be re-established and connected to its present time.
Likewise, we have seen some personal accounts from patients on how schizophrenia causes the sensation that time has stopped. However, this is not restricted to only those with mental illness. For instance, according to Wilhelm Lange-Eichbaum (1946), a German psychiatrist, Nietzsche experienced a timeless moment as a result of his illness. Nietzsche himself wrote about the slowing down of his time sense as a result of his indulging in some of his feelings. Lange-Eichbaum42 was also persuaded that Nietzsche undoubtedly exhibited schizophrenia-like symptoms. However, Nietzsche’s experience with schizophrenia might have intensified this feeling of timelessness and gave him ‘Übermensch’ insight, assisting him in writing his ‘Also sprach Zarathustra’ and made him immortal.
In the modern era, a lot of research has been conducted on the timing inefficiency of patients with schizophrenia. In an experiment done by Butler et al,43 they showed the inability of those with schizophrenia to appropriately coordinate events in time. Most patients’ dysfunctional coordination of events in time exhibits these problems: lengthened windows of simultaneity, shifted integration windows,44 impaired two-pulse resolution,45 reduced sensitivity in temporal contrast detection46 47 and lowered thresholds in flicker-fusion detection.48
If we take rhythmic music as time, we observe a very strange behaviour among these patients: their movements become wooden and mechanical like a robot or doll, and it looks like they have lost the former naturalness of their movements. In other words, they become strangely ‘unmusicked’. To an observer, they are like a still photo, a frozen frame which is stuck in a motionless moment—in a statelessness and timelessness of unreality. We also see in Nietzsche’s descriptions of schizophrenia a ‘degeneration of a sense of rhythm’.
Having observed schizophrenia as a deficiency in precise timing, one may suppose that this illness does not come from one singular cause. Instead, it arises from a number of increasing dysregulations, and a destabilising system which can be understood from multiple angles (here, the origin of this dysregulation from the concept of time perspective was mentioned). According to the principle of reafference and the importance of temporal integration between the CNS and the outside world, schizophrenia can be a dysregulation of precise timing or a range of dysregulations within a complex system. When viewed this way, aetiologies of the illness can come out at many points, as can sources of healing. This perspective towards schizophrenia might also give clues to a better healing system for this mysterious illness.
I close this article with the main theme from the book by Louis Sass, Madness and Modernism (1992). In his book,49 he relates schizophrenia to the artistic history of the past century—emphasising that the ‘descriptive’ psychopathology of schizophrenia is still a lucrative and influential collection of information for all those interested in the mechanisms of the mind for ‘normals’, as well as for those attempting to understand the nature and causes of schizophrenia.
Ethics statements
Patient consent for publication
Ethics approval
Not applicable.
References
Morteza Izadifar obtained a master’s degree of Cognitive Studies, Psychology, and Neuroscience in Europe. After his graduation, he began a journey as a research traveler around Europe. He has worked as a research fellow in different cognitive neuroscience labs in Spain, France, Italy and Germany. He obtained his doctorate at the Institute of Medical Psychology, the Ludwig-Maximilians-Universität München (LMU) in Munich, Germany where he studied the underlying neuronal mechanisms of functions of the brain such as subjective time perception, time and timing of the brain, time perception in imagery, cognitive neuroscience of imagery, discrete and continuous flow of consciousness, neural foundations of creativity, multisensory integration and schizophrenia. He is interested in translating research findings into clinical practice and is committed to helping people benefit from the many advances of psychological sciences. He is now doing postdoctoral training collaboratively with his previous institute (the Institute of Medical Psychology, Germany) and the University of Arizona, USA (Department of Psychology and Neuroscience).

Footnotes
Contributors MI is the main author of the manuscript.
Funding The authors have not declared a specific grant for this research from any funding agency in the public, commercial or not-for-profit sectors.
Competing interests None declared.
Provenance and peer review Not commissioned; externally peer reviewed.