Article Text

Abstract
Behavioural inhibition is a biologically based risk factor for anxiety disorders. Children with behavioural inhibition are shy, cautious and avoidant of new situations. Much research on behavioural inhibition has focused on the amygdala as an underlying neural substrate and has identified differences in amygdala function and volume; however, amygdala findings have yet to lead to meaningful interventions for prevention or treatment of anxiety disorders. The bed nucleus of the stria terminalis (BNST) is a prime candidate to be a neural substrate of behavioural inhibition, given current evidence of BNST function and development in human research and animal models. Children with behavioural inhibition have an increased startle response to safety cues and an increased cortisol response to social evaluative situations, both of which are mediated by the BNST. In rodents, activation of the BNST underlies contextual fear responses and responses to uncertain and sustained threat. Non-human primates with anxious temperament (the macaque equivalent of behavioural inhibition) have increased BNST activity to ambiguous social situations, and activity of the BNST in anxious temperament is significantly heritable. Importantly, the BNST is sexually dimorphic and continues to develop into adulthood, paralleling the development of anxiety disorders in humans. Together, these findings suggest that further investigation of the BNST in behavioural inhibition is necessary and may lead to new avenues for the prevention and treatment of anxiety disorders.
- anxiety disorders
- fear response
- behavioural inhibition
- bed nucleus of the stria terminalis (BNST)
- amygdala
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
- anxiety disorders
- fear response
- behavioural inhibition
- bed nucleus of the stria terminalis (BNST)
- amygdala
Anxiety disorders
Anxiety disorders are chronic,1 highly impairing2–4 and associated with the development of comorbid mood and substance use disorders.5–7 Individuals with anxiety disorders rarely seek treatment (approximately one in five individuals with an anxiety disorder seek treatment in both the USA and China)8 9 and responses to current treatments are modest at best.10 11 The estimated cost of anxiety disorders in America alone is over US$45 billion.12 13 Given this substantial societal burden of anxiety disorders, more work is needed to prevent and treat these devastating diseases. Understanding the neurobiology of risk for anxiety disorders is a critical first step.
Behavioural inhibition
Anxiety disorders are often preceded by behavioural inhibition,14–17 the tendency to be shy, cautious and avoidant of novelty.18 This temperament begins in infancy as high reactivity to novelty. Infants who are highly reactive to novel stimuli and distressed on separation from their mothers typically become behaviourally inhibited children.18–20 Behavioural inhibition represents the most extreme 15% on a continuum of reactivity to novelty and many, but not all, children with behavioural inhibition develop anxiety disorders later in life.15 Behaviourally inhibited children most commonly develop social anxiety disorder; one meta-analysis found that 43% of behaviourally inhibited children develop social anxiety disorder by late childhood or early adolescence.15 However, childhood behavioural inhibition also increases risk for other anxiety disorders, including generalised anxiety disorder,21 22 phobias,22–24 and agoraphobia.23 Childhood behavioural inhibition also increases risk for a number of other known comorbidities of anxiety disorders, including depression22 25–27 and substance use disorders.28 29
Behavioural inhibition can be observed across cultures30 31 and is evolutionarily conserved across species, including earthworms, octopuses, non-human primates and others (for a review, see Gosling and John32). Additionally, behavioural inhibition is heritable33 34 and has moderate stability across the lifetime;18 35 taken together, these data suggest that behavioural inhibition has a biological basis. Because behavioural inhibition precedes the onset of anxiety disorders, has a known biological basis and is measurable, understanding the neurobiology of behavioural inhibition may allow us to gain traction on the prevention and treatment of anxiety disorders.
In 1988, Dr Jerome Kagan proposed that the biological bases of behavioural inhibition were hyperactivity of the amygdala and hyper-reactivity of the sympathetic nervous system; these changes led to increased reactivity to novelty.36 Physiological studies in children with behavioural inhibition have shown that behavioural inhibition is associated with decreased threshold for activation of the sympathetic nervous system,37 less parasympathetic input, as measured by a higher and more stable (less variable) heart rate,36 38–40 and an increased sympathetic response, as measured by increased heart rate reactivity to stressors,36 41 larger pupillary diameter, increased laryngeal muscle tension, and increased urinary norepinephrine concentration.36 39 Cortisol and related hormones (ie, corticotropin-releasing hormone (CRH)) modulate these stress responses (both initiation of the stress response and inhibition through a negative feedback pathway)42 and much of the early research into the biological basis of behavioural inhibition focused on the cortisol response.
Several studies of the hypothalamic-pituitary-adrenal (HPA) axis response in behavioural inhibition have found that behavioural inhibition is associated with higher cortisol concentrations at baseline and in response to stress. Inhibited children had significantly higher cortisol concentrations at home and in the laboratory39 41 43. Pérez-Edgar et al44 found that behavioural inhibition was associated with higher morning salivary cortisol and that increased cortisol concentration and increased negative affect predicted more social withdrawal behaviour, a precursor of social anxiety. Further investigation showed that increased cortisol concentration in children aged 4 years was significantly associated with increased social withdrawal behaviour, specifically in boys with a history of high negative affect. High cortisol might sustain negative affect and behavioural inhibition in boys with high negative affect early in life. Children with behavioural inhibition may be more sensitive to social stressors, resulting in increased cortisol reactivity—this increased sensitivity to social stressors may be governed by alterations in amygdala, hippocampus and bed nucleus of the stria terminalis (BNST) response.42
Another consistent finding in individuals with behavioural inhibition is increased startle reactivity. In response to a sudden and intense stimulus, humans and lower mammals display a sudden eyeblink response.45 Infants who were highly reactive to novelty had an increase in startle amplitude when a stranger approached.43 Children with behavioural inhibition have a shorter latency to startle response46; children with behavioural inhibition who had a larger startle response during safety developed more internalising problems and social anxiety 2 years later.47 Adolescents with behavioural inhibition who met criteria for an anxiety disorder had heightened startle to safety cues compared with behaviourally inhibited adolescents without an anxiety disorder, but there were no group differences in startle to fear cues.48 Additionally, Schmidt et al found no temperament differences in magnitude of fear-potentiated startle in young children.41 These findings suggest that children with behavioural inhibition have elevated startle to safety, but not fear cues, and that heightened startle to safety cues may predict risk for the development of anxiety disorders. Fear-potentiated startle (ie, increased startle after seeing a fearful stimulus) is thought to be mediated by the amygdala, whereas anxiety-potentiated startle (ie, increased startle in a context that might be associated with threat) is thought to be mediated by the BNST.49 While safety cues signal the lack of threat in these paradigms, behaviourally inhibited children may interpret the entire startle paradigm as anxiety-provoking and thus have elevated startle to safety cues mediated by BNST activation; however, that hypothesis has yet to be tested at this time. Consistent findings of elevated startle to safety cues in behavioural inhibition suggest that BNST function might be altered in behavioural inhibition.
Neurobiology of behavioural inhibition
A number of recent manuscripts have reviewed the neurobiological underpinnings of behavioural inhibition.50–52 Inhibited temperament was initially described as the tendency to avoid and be wary of novel people, places or things. As the amygdala responds to novel, fearful and salient stimuli in the environment53 54 and has efferent projections to the limbic system, sensory cortex, and brainstem,53–56 amygdala hyperactivity was first proposed as the neurobiological basis of inhibited temperament.39 57
Early neuroimaging studies found that behavioural inhibition was associated with a faster,58 larger50 59 and more sustained58 amygdala response to novel social stimuli. Behavioural inhibition was also associated with increased amygdala response to newly familiar social stimuli60 61 and internal reflection about social stimuli.62 Consistent with evidence in rodents that sustained stress and increased cortisol activity leads to larger amygdala volume,63–65 amygdala volume and activation are increased in young adults with behavioural inhibition.66 Despite evidence for amygdala hyperactivity in behavioural inhibition, identifying these changes in amygdala function and structure has not led to differences in treatment or prevention of anxiety disorders. Failure of amygdala findings to lead to changes in prevention or treatment suggests that we should search for additional neurobiological markers of behavioural inhibition.
Evidence for the BNST in anxiety
Rodent models
The BNST is a small structure in the medial forebrain which is considered to be part of the ‘extended amygdala’ (figure 1). The BNST and amygdala have similar connectivity and gene expression profiles67 (figures 2 and 3). In both rodents and humans, there is an increasing startle response from neutral context, to conditioned stimulus, and highest in the contextual fear context; in humans, subjective anxiety also increases from neutral context to predictable threat, to peak at the unpredictable threat context.68 Rodents who experience threat in one context then go on to avoid that context.69 Increased contextual avoidance in rodents parallels symptoms seen in anxiety disorders and behavioural inhibition (table 1). Alterations in BNST function may underlie the behavioural and physiological changes associated with behavioural inhibition and provide new avenues for treatment and prevention of anxiety disorders. Other recent reviews70 71 have described emerging research in the BNST and provide an excellent foundation about this brain region; this review specifically focuses on the relationship of the BNST to developing anxiety disorders.
Relationship of anxiety disorders to BNST characteristics

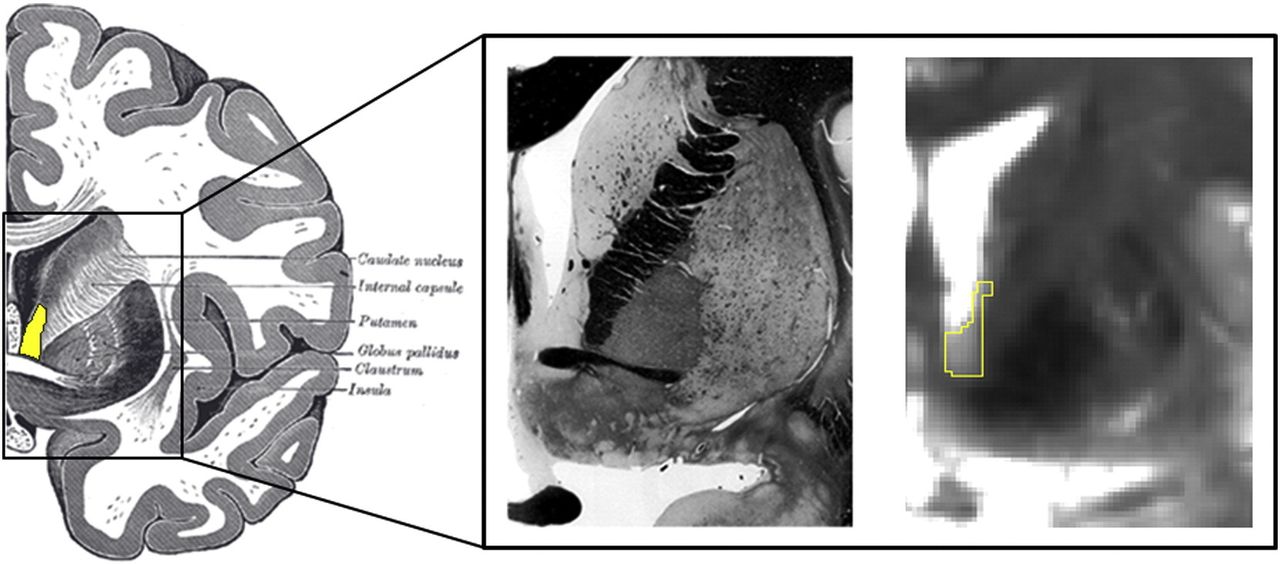
The human bed nucleus of the stria terminalis (BNST). (Left) Human illustration with BNST highlighted in yellow. (Middle) Human fixed tissue slice. (Right) BNST mask on a 7T gradient spin echo MRI image. Used with permission from Avery et al77.

Rodent BNST connectivity. The rodent BNST is connected to a number of limbic, subcortical and cortical regions. Figure adapted from Sokolowski and Corbin96. BNST, bed nucleus of the stria terminalis; Hipp, hippocampus; NST, nucleus tractus solitarious; NuAc, nucleus accumbens; PAG, periaqueductal gray; PB, parabrachial nucleus; SI, substantia innominata; VTA, ventral tegmental area.

{kind=link}
{kind=link}
{kind=link}
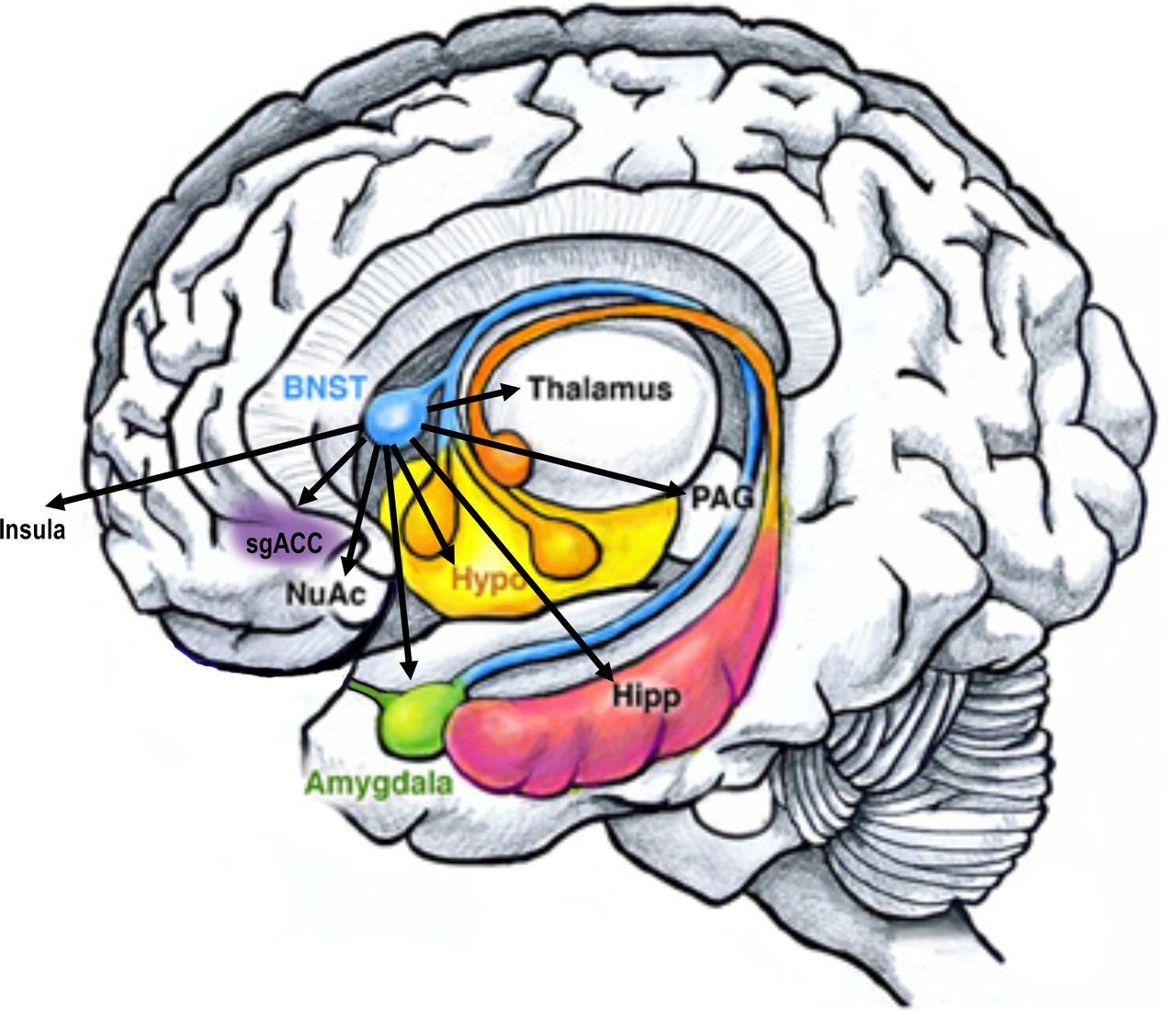
Human BNST connectivity. The human BNST has been shown to be functionally and structurally connected to similar regions as the rodent BNST, and to additional cortical regions. Adapted from Sokolowski and Corbin96. BNST, bed nucleus of the stria terminalis; Hipp, hippocampus; NuAc, nucleus accumbens; PAG, periaqueductal gray; sgACC, subgenual anterior cingualte cortex.
Decades of research have demonstrated that while the amygdala responds to certain threat or fear stimuli, the BNST responds to uncertainty and sustained threat.49 This dichotomy was initially discovered in rodent models using the startle response. Startle magnitude in rodents is larger in bright light (light-enhanced startle; equivalent to darkness for humans) and after hearing a cue previously paired with a shock (fear-potentiated startle)45 (table 1). BNST lesions reduce the light-enhanced startle response, but do not affect the fear-potentiated startle response.45 Infusion of CRH into the BNST increases the magnitude of both fear-potentiated startle72 and startle in response to contextual fear (environment in which shock has occurred).69 Rodents with BNST lesions have lower conditioned fear responses to long-duration conditioned stimuli (10 min between conditioned stimulus and unconditioned stimulus), which may represent a period of persistent anxiety or uncertain threat.73
Other phenotypes associated with behavioural inhibition have also been shown in rodent models to be related to BNST function. BNST lesions decrease latency to enter and increase exploration time in the open arm of an elevated zero maze, another model of avoidance of novelty and anxiety in rodents.73 Social defeat (ie, an animal’s failure to continue to defend itself after being defeated by a rival) is mediated by CRH activity in the BNST74 and animals experiencing inescapable shock do not give up escaping their cage if they have BNST lesions.49 Mounting evidence in rodent models suggests that BNST activity underlies many of the symptoms seen in anxiety disorders in humans.
Human neuroimaging
In humans, activation of the BNST has been provoked using virtual reality contexts (table 2). In pioneering work by 75 76Alvarez et al 75 and Torrisi et al 76 virtual reality contexts have been used to represent certain threat (in which an auditory cue always predicted a shock), uncertain threat (tones and shocks were presented in an unrelated fashion), and neutral contexts (in which tones were presented, but there were no shocks). These virtual reality paradigms can be used inside the functional MRI scanner to replicate predictable and unpredictable threat in rodent paradigms. Prior to the start of the MRI scan, participants were shown each context and instructed on the context-threat pairing. They were instructed that in one context (ie, a bank), the tone would be paired with a shock, in a second context (ie, a casino), the tone and shock would have no relationship, and in a third context (ie, a restaurant), they would hear the tone, but there would be no shock. Between the indoor contexts, participants viewed an outdoor context that represented the intertrial interval. Subjective anxiety increased linearly from neutral context, to the predictable threat context, to the unpredictable context.75 Replicating findings in rodents, the predictable threat context activated the amygdala, whereas the unpredictable threat context activated the BNST.75
Critical studies of human and non-human primate BNST function and connectivity related to behavioural inhibition and anxiety disorders
There are a number of methodological challenges in examining BNST differences with functional MRI in humans. The BNST is a small structure (~1/10th the size of the amygdala) located in the basomedial forebrain and is difficult to delineate using conventional imaging methods (figure 1). Recent advances in neuroimaging have allowed for smaller voxel sizes and thus more accurate imaging of the BNST75 77–79 and the BNST has been reliably identified in 3 Tesla MRI scans.80
Functional neuroimaging studies of the BNST have demonstrated similar findings in rodents and non-human primates (table 2). The BNST has both structural and functional connectivity with limbic regions, including the amygdala and hippocampus, the basal ganglia, including nucleus accumbens, caudate and globus pallidus, as well as the thalamus (figure 3).77 81 Additionally, the BNST has significant functional connectivity with the medial prefrontal cortex and insula.77 Recently, BNST functional connectivity with the hypothalamus, sublenticular extended amygdala and periaqueductal gray has been demonstrated using ultra high-field functional MRI.81
Non-human primate models
Kalin et al have examined anxious temperament, a non-human primate analogue of behavioural inhibition. Anxious temperament is defined as the triad of increased freezing, decreased cooing response, and increased cortisol in response to a non-threatening intruder (unfamiliar human displaying a profile to the macaque and not making eye contact, known as the ‘no eye contact’ (NEC) paradigm).82 83 During the NEC paradigm, the intruder presents an ambiguous potential threat, as the intruder’s intentions are not known based on the lack of eye contact. Non-human primate models have several advantages in the study of the neurobiology of anxiety risk. Non-human primate models allow for precisely characterised genetics, mechanistic studies, including precise brain lesion studies and the ability to repeatedly conduct positron emission tomography imaging over time.
Early studies showed that anxious temperament and increased freezing during the NEC paradigm were associated with significantly increased glucose uptake in the BNST (table 2).82 84 Additionally, increased serotonin transporter availability in the BNST was associated with anxious temperament and increased glucose uptake during the NEC paradigm.85 In a large-scale study examining the heritability of brain activation in anxious temperament, Oler et al86 found that anxious temperament-related activation in the amygdala, insula and BNST was significantly heritable and activation in the BNST during a social task was the most heritable of the brain regions identified. Activation of the orbitofrontal cortex may be a critical component of freezing in anxious temperament. Macaques with orbitofrontal cortex lesions did not display increased freezing in response to the NEC intruder paradigm and did not have increases in BNST activation, suggesting that orbitofrontal cortex influences on the BNST are critical for full expression of the anxious temperament phenotype.87
Human behavioural inhibition
Neuroimaging studies of BNST in humans with behavioural inhibition have been limited. To date, one study in humans with trait social anxiety88 showed that across all subjects, the BNST activates to social stimuli following a positive or negative cue (ie, unpredictable valence). Individuals with higher social anxiety had an increased BNST response to unpredictable images. Individuals with more social anxiety also had increased connectivity between the BNST with both the posterior cingulate cortex and ventromedial prefrontal cortex when viewing unpredictable threat images. This study provides preliminary evidence for a role of the BNST in a phenotype similar to behavioural inhibition. Tasks that activate the BNST, such as those that evoke uncertainty and potential threat, should be used in inhibited temperament to test for differences in BNST activation.
Future directions
The BNST does not function in isolation and a number of recent studies have examined connectivity of the BNST in humans.76 77 89 BNST connectivity is likely to also be altered in behavioural inhibition and patterns of change in BNST connectivity may shed light on anxiety risk. Applying virtual reality paradigms such as those in the study by Grillon et al75 76 may be helpful in understanding differences in BNST neurocircuitry in behavioural inhibition.
The BNST is sexually dimorphic90 and sex hormones can alter BNST structure and function.49 Given that anxiety disorders are more common in females,7 alterations in BNST structure and response to sex hormones may mediate some of these gender differences. Sexual differentiation of the BNST in humans may extend into adulthood.91 As the BNST continues to develop throughout puberty, understanding these changes in BNST development may also help us to understand anxiety disorder risk, as many anxiety disorders have their onset during puberty.
Summary
A number of factors point to the BNST as a critical mediator of behavioural inhibition and anxiety risk (see tables 1 and 2), including its relationship to contextual fear and anxiety, cortisol response and hyperstartle. Understanding alterations in BNST response may lead to new avenues for treatment and prevention of anxiety disorders. Exploring the development of BNST over time in human subjects and changes in the patterns of BNST connectivity may provide critical information regarding the development of anxiety disorders.
Acknowledgments
The author would like to thank Dr Philip Cawkwell for helpful feedback on the manuscript. The author would like to thank Dr Jennifer Blackford and Dr Suzanne Avery for their helpful discussions about the BNST and behavioural inhibition.
References
Jacqueline Clauss, M.D, Ph.D., obtained her doctoral degrees from Vanderbilt University and currently serves as a child and adolescent psychiatry fellow at Massachusetts General Hospital/McLean Hospital. Dr. Clauss is an expert in the neurobiological mechanisms implicated in the development of psychiatric disorders, with focuses on building resilience in high risk children and using neuroimaging to understand underlying neurobiology.

Footnotes
Contributors JC developed the ideas, wrote the manuscript and edited the manuscript. JC conceived and wrote the current manuscript.
Funding JC is supported by the MGH/McLean Research Concentration Program (NIMH R25‐MH094612) and by NIMH 3-R01-MH109562-04S1.
Competing interests None declared.
Patient consent for publication Not required.
Provenance and peer review Commissioned; externally peer reviewed.